Phototransduction
This line of research was done in collaboration with Christopher W. Tyler (Smith-Kettlewell), Daniel Tranchina (NYU Courant Institute), Trevor D. Lamb (Eccles Institute of Neuroscience, Australian National University), Paul A. Liebman (Department of Biochemistry & Biophysics, School of Medicine, University of Pennsylvania) and Spero C. Nicholas (Smith-Kettlewell).
<><><>
Phototransduction (PTD) is the process by which rod and cone photoreceptors convert light energy into bioelectric signals that eventually leave the retina and initiate responses in the brain that we experience as “seeing” (vision). Between 1995 and 2006 my colleagues and I developed detailed models of the biochemistry and biophysics of phototransduction.
Phototransduction in the Context of Perception & Cognition
It is remarkable to consider that the stimulus for vision – light falling on the retina – has only 2 dimensions: intensity (number of photons arriving at the retina per unit of time) and wavelength composition (which we perceive as the different color of the rainbow). And yet the phenomenology of vision, what we actually see in visual scenes, has many perceptual dimensions: brightness, color-hue, color-saturation, contrast (relative perceived brightness between objects or scene elements), size, location (relative to you), depth (perceived distance of an object) and relative depth (as in figure-ground segregation, discerning “what is in front of what” in any given scene), visual texture. Moreover, we can visually discern more abstract elements and qualities that intersect with our other senses, such as wetness/dryness, temperature (red-hot coals, a blue flame), and with our higher level cognitive-perceptual experience, such as perceiving other’s emotions, perceiving danger, and ultimately even leading to even more complex experiences like beauty and the many varied aesthetic visual experiences.
Thus, the scientific project of understanding how light energy is transformed into bioelectric signals that lead to the vast multidimensional visual experience described above is foundational. The processes that initiate vision and which permit creatures to live and evolve across generations are thus profoundly important: understanding PTD and how our sensory receptors can function over a huge dynamic range of 10 billion to one is a deeply important question that has fascinated visual neuroscientists for almost a century.
The Vertebrate Eye & Retina

The vertebrate eye, and specifically the structure and function of the retina, stands as a magnificent exemplar of the biological complexity and physiological adaptability that can result from eons of natural selection. It is noteworthy that the “design” of the retina – the number and types of cells, their structure and function and connectivity – is, in large measure, the same across all species in the subphylum Vertebrata. And the “design” of the retina has been preserved for over 500 million years.
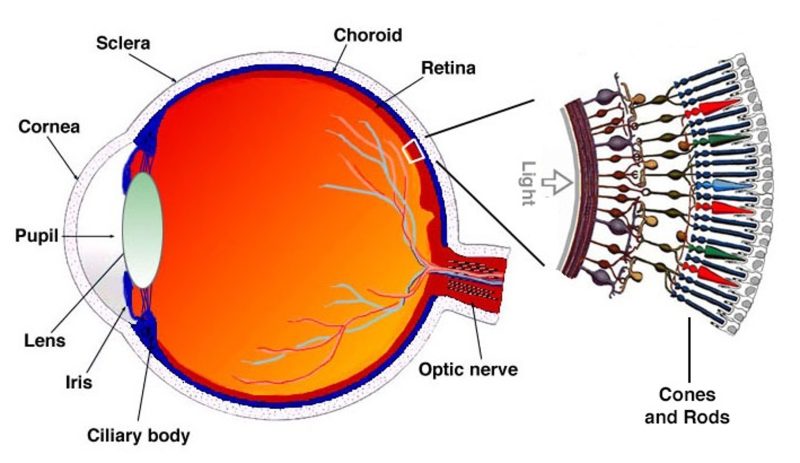

LEFT: A schematic depiction of the anatomy of the eye showing the optical componenets (cornea, pupil, lens) and a blown-up “cartoon” depiction of a piece of retina with the photoreceptors localized at the back of the retina, facing away from the light, near their blood supply. RIGHT: An elegant, detailed drawing from 1887 of an actual vertebrate retina dissected from an animal cadaver by the Italian anatomist Ferruccio Tartuferi (1852-1925). Tartuferi painstakingly drew all the retinal cells and layers he observed. His magnificent drawing is impressively close to modern micro-anatomical images of the vertebrate retina. Light would enter the retina from the left of the drawing, with the photoreceptors, rods and cones on the right, in the back of the retina.
Rod photoreceptors, which much more sensitive than cones, actually evolved after cones. Unlike cones, rods are able to signal the absorption of single photons of light and thus are the receptors we vertebrates depend on to function in very low light levels. The need to survive the dangers of the night are clearly paramount, and it has been argued that our exquisite rod sensitivity is responsible for the very survival of the vertebrate line.
How rods are able to reliably signal the absorption of single photons of light is a scientific mystery still under investigation. Some of my research in collaboration with my colleagues addressed this very mystery.
Modeling Phototransduction
My colleagues and I developed and refined a model of rod phototransduction that was able to reproduce the remarkable ability of rods to reliably signal the capture of a single photon of light. We knew as early as 1942 that it was quite likely that humans could detect the absorption of very few (1-5) photons by as many rods (Hecht, Shlaer,& Pirenne. 1942. Energy, quanta, and vision. Journal of General Physiology, 25, 819 840. https://doi.org/10.1085/jgp.25.6.819)
The first physiological demonstration of this ability in rods awaited the elegant recording techniques in the lab of Denis Baylor in the late 1970s. How rods could achieve this fidelity of response was a scientific mystery since each of the biochemical reactions elicited by the capture of one photon by one rhodopsin molecule (R) in one rod photoreceptor, where each rod cell contained 10 million (or more) R’s, were inherently stochastic (random).
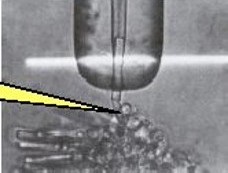
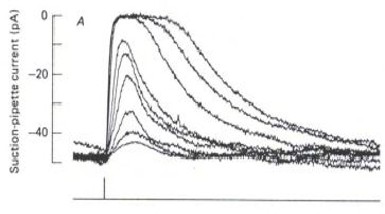
LEFT: An outer segment (OS) of a single rod photoreceptor is drawn into a glass suction electrode (large vertical object). A 10 ms flash of light (the white bar) is projected onto the OS while the rod’s electrical response is recorded via a microelectrode (illustrated in yellow). A series of flashes of increasing intensity elicit the family of electrical responses shown on the RIGHT. (from the lab of Denis Baylor & colleagues, c. 1979)
In our computational model (Hamer et al.; 2003, 2005), we implemented the most well-establiahed biochemical events after a molecule of R captured a photon and was converted into an active enzyme, R*. Based on prior research in the field, we hypothesized that an R* deactivated back to R in a sequence of steps facilitated by sequential phosphorylation (Pn) of various sites on the large R* molecule. There were at least 6 and possibly 7 known phosphorylation sites. Our model showed that if successive Pn of these sites led to progressive inactivation of R* back to R, the remarkable reproducibility of the single-photon response of rods could be accounted for quantitatively. Moreover, our model could reproduce the physiological responses of rods across an enormous range of light levels, from single-photon regime to highly light-adapted regime where rods were responding to absorption of many thousands of photons per second.

Comparison of various features of rod responses (LEFT column) to absorption of very few photons (0,1,2..etc) to our model predictions (RIGHT column) for a series of experimental manipulations.(Hamer et al. 2005). The top pair of graphs show responses to absorption of one (red) or two (or more, green): gray curves represent “failures”, i.e., when the stimlus failed to evoke any response: empiral data (left) and our model responses (right) are shown.
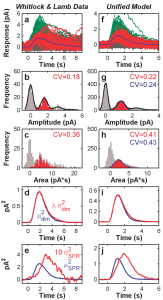
Our model was even able to reproduce the responses of rods in experimental animals whose biochemical phototransduction “machinery” had been genetically altered so as to selectively alter (or silence) specific reactions in the phototransduction sequence.

LEFT: Single-photon responses in rods that had been genetically altered to selectively eliminate 4 critical biochemical reactions in the phototransduction sequence. The bottom black curve shows the response from normal rods RIGHT: Our model responses when the same biochemical steps were eliminated from the model. (from Hamer et al., 2005).)
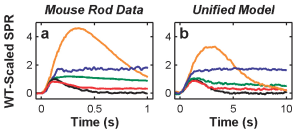
Remarkably, our model was able to account for this large range of responses, including the responses from the genetically altered rods, without changing any parameters in the model: The physiological responses that our model could reproduce were: “(1) SPR waveforms and statistics (as in Hamer et al., 2003); (2) a full dark-adapted flash-response series, from dim flash to saturating, bright flash levels, from a toad rod; (3) steady-state light-adapted (LA) responses, including LA circulating current (as in Koutalos et al., 1995) and LA flash sensitivity measured in rods from four species; (4) step responses from newt rods (Forti et al., 1989) over a large dynamic range; (5) dynamic LA responses, such as the step-flash paradigm of Fain et al. (1989)), and the two-flash paradigm of Murnick and Lamb (1996); and (6) the salient response features from four knockout rod preparations. The model was able to meet this stringent test, accounting for almost all the salient qualitative, and many quantitative features, of the responses across this broad array of stimulus conditions, including SPR reproducibility.” (see Hamer et al, 2005).
The contribution of our model to understanding how photoreceptors can reliably signal absorption of very few photons and also respond to an enormous range of light intensities was discussed in “The Retina & Vision”, Chapter 19 in Vol. II of the 2nd Edition of Keener & Sneyd’s, Mathematical Physiology II: Systems Physiology (Interdisciplinary Applied Mathematics, 8/2) (2008), Springer:
“…One of the most striking features of phototransduction is that not only can a rod respond to a single photon, it does so in a reproducible way that varies little from photon to photon. At first glance this might seem uninteresting, until one realizes that a single photon response is mediated by a single rhodopsin molecule. Since the behavior of a single molecule must, necessarily, be stochastic in nature, how can this produce a reproducible single-photon response? Baylor et al. (1979) were the first to study in detail this question, which has long been considered one of the most puzzling and important problems in photoreceptor physiology…”
As shown above, our multiple-phosphorylation model was able to account for the observed statstics of photoreceptor resonses at very low light levels, but a comprehensive model must be also able to account for the photoreceptor’s behavior at high light levels spanning the cell’s full physiological dynamic range. On this point, Keener & Sneyd elaborate in their 2026 Third Edition of “Mathematical Physiology“:
“Although multiple phosphorylation steps provides a plausible answer to how a single photon can generate a reproducible response, to understand how phototransduction works at all light levels it is necessary to construct a single model that can reproduce not only the single-photon response, but also the responses to flashes and steps of light, both bright and dim. This constitutes a severe set of tests against which to validate any model of phototransduction, and it was not until 2005 that such a model appeared (Hamer et al. 2005). Hamer et al elaborated their 2003 model and were able to capture many qualitative and quantitative features of vertebrate rod responses from several species. Using a single set of parameters (except for sensitivity adjustments to account for differences between species and experimental preparations), Hamer et al. were able to account for (among other things): (i) single-photon responses and reproducibility statistics, (ii) dark-adapted flash responses over a wide range of flash intensities, (iii) step responses over a 4.7 log unit range, and (iv) steady-state light-adapted responses under normal conditions and when Ca2+ feedback was prevented.” (p. 1060)
Analysis of Ca++-dependent Gain Changes in PDE Activation in Vertebrate Rod Phototransduction (2000) Download
Computational Analysis of Vertebrate Phototransduction: Combined Quantitative and Qualitative Modeling of Dark- and Light-Adapted Responses in Amphibian Rods (2000) Download
Multiple Steps of Phosphorylation of Activated Rhodopsin Can Account for the Reproducibility of Vertebrate Rod Single-photon Responses (2003) Download
Toward a Unified Model of Vertebrate Rod Phototransduction (2005) Download