Somatosensory: Sensory Detection Via Receptors in the Skin
My Ph.D. research focused on vibration sensitivity of the skin. Specifically, in my experiments, vibratory stimuli were applied to the skin whose vibration frequency and spatial parameters were chosen to selectively activate, and be detected by, different mechanoreceptor classes in the skin: either by the extremely sensitive, and very rapidly adapting Pacinian Corpuscles (P), or other slower, less sensitive non-Pacinian (NP) classes of receptor.
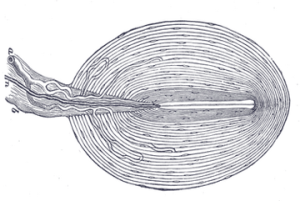
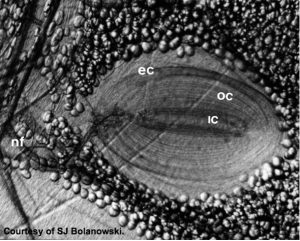
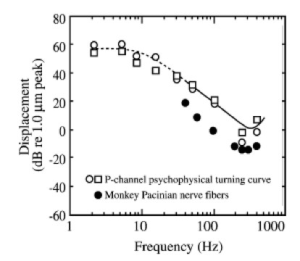
LEFT: Detailed sketch of a Pacinian Corpuscle (approx 1 mm diam). The mechanical transduction occurs at the nerve in the cemter of the many layers of the capsule that acts as a mechanical bandpass filter with a maximum transmission of vibratory stimuli at approximately 200-300 Hz. The actual mechanotransduction occurs at the center where the specialized mechanosensitive nerve ending is. CENTER: An actual photomicrograph of a Pacinian Corpusclet (Courtesy of S. Bolanowski). RIGHT: Temporal frequency tuning of individual primate Pacinian corpuscles (filled circles) compared with the psychophysical tuning (vibration amplitude thresholds vs temporal frequency for sinusoidal vibration stimuli) for human subjects. Both the primate physiology and human psychophysics have the same temporal tuning with minimum vibration thresholds occurring at around 200 Hz. Both have minimum threshold amplitudes of between 0 and -10 dB re: 1 micron.
The P receptors have a bandpass sensitivity profile with maximum sensitivity for vibration frequencies of about 200-400 Hz. The NP receptors were most sensitive to much lower vibration frequencies (approx. 20-30 Hz), and were very much less sensitive (higher psychophysical thresholds).
I found that when spatial and temporal frequencies were carefully chosen, the P and NP receptor pathways could be almost independently activated, with psychophysical (perceptual) detection, even in the presence of vibratory masking stimuli, being determined mainly by only one or the other sensory channel.
My research demonstrated that humans could reliably detect vibrations applied to the glabrous (non-hairy) skin of the hand at extremely low vibration amplitudes when the stimuli were designed to activate the P receptors. Moreover, in my 1979 Ph.D. Dissertation, I presented data showing a striking facilitative effect of a masking stimulus that enhanced rather than impeded detection of a test stimulus at low masker levels. This masking profile came to be known as “Dipper Function“. The phenomenon itself (termed negative masking or pedestal effect) was first noted in audition and then later in visual masking studies (e.g., Nachmias & Kocher, 1974;Legge & Foley, 1980). Remarkably, in vibrotactile masking, the detectability of a vibration increment was maximally enhanced at very low masker amplitudes, amplitudes even below the threshold for detection of the masker alone. Note the data points below zero on the ordinate in the graph below reach a minimum (maximum negative masking) below the threshold for masker vibration (zero on the abscissa), that is even before the masker itself is detectible.
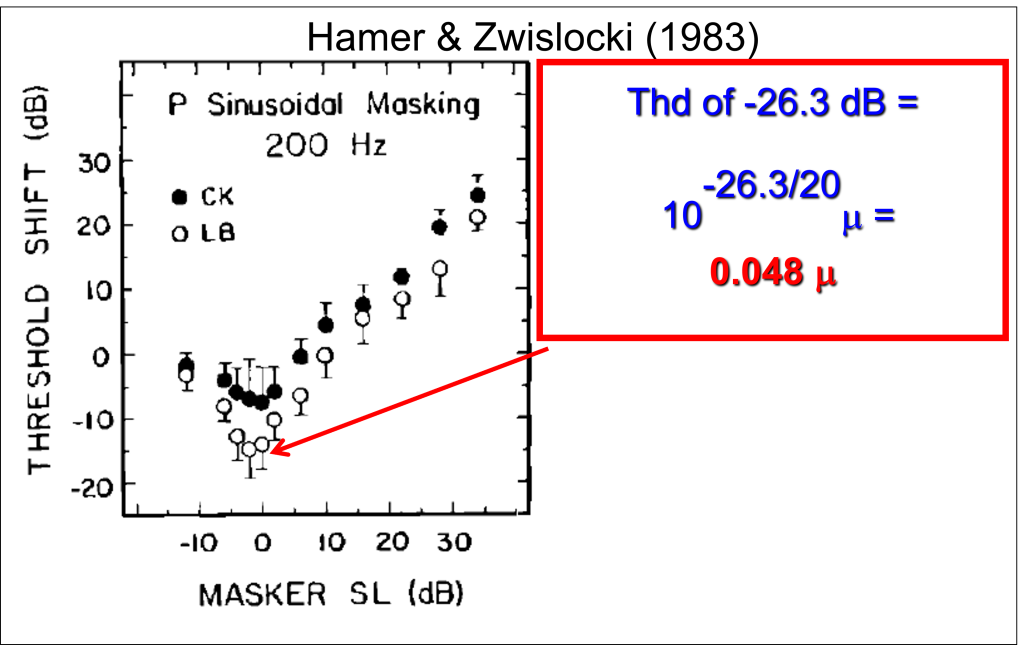
For some subjects, the threshold for detection of a brief, P-selective vibration increment in the presence of a very low-amplitude background masking vibration was extraordinarily low. For subject LB, the masking vibration enhanced his sensitivity by about 15 dB, yielding an absolute calibrated threshold vibration amplitude of -26.3 dB re: 1 micron (see figure above). LB could reliably detect 200 Hz vibrations with amplitudes of 0.048 microns. This is about 10 times smaller than the wavelength of blue light, or 1000 times smaller than the diameter of an average human hair. This remarkable sensitivity to high-frequency vibrations of the skin where Pacinian Corpuscles are abundant (such as the glabrous skin of the hands and feet) provides us with exquisite sensitivity to tactile textures as we explore surfaces with our hands (which have the highest density of Pacinian corpuscles in all of our skin surface).
Note in the graph above that the minima (lowest threshold for detecting the stimulus in the presence of a background masker) ocurred below zero on the abscissa. My analyses showed unequicocally that all the data points below zero on the abscissa could be precisely predicted by a model that assumed (1) the Pacinian channel had a hard threshold with little or no spontaneous activity in the abscence of a stimulus and (2) that that low-amplitude maskers were simply acting as pedestals: as a subthreshold pedestal (masker) amplitude increased, the extra vibration amplitude needed to exceed the Pacinian threshold decreased. Hence, all the masking data beow zero on the abscissa could be modeled with one parameter: a hard (noiseless) threshold.
Vibrotactile Masking of Pacinian and non-Pacinian Channels (1982) Download